在路径模型中寻找守恒量
此示例显示如何使用SBIocons部分函数用于在SimBiology®模型中查找守恒量。
目标
使用
SBIocons部分确定两种糖酵解模型中的保守量t . brucei.在分析这些模型时使用计算出的守恒量。
背景
布氏锥虫是导致非洲昏睡病的单细胞真核寄生虫。这种生物通过从宿主血液中代谢葡萄糖而在受感染的宿主体内生存。在t . brucei与其他锥虫一样,糖酵解的很大一部分发生在称为糖体的特殊细胞器内。
为了研究糖体的功能,Bakker等人(20001997)构建并验证了糖酵解的计算模型t . brucei这明确包括糖体区隔。他们将该模型的性质与不含糖体的衍生模型的性质进行了比较。在其他结果中,他们发现在缺少糖体的情况下,糖酵解途径中的磷酸己糖中间产物可以积累到对细胞有害的高水平。在他们的分析中,Bakker等人能够解释糖体提供的代谢物的分区如何防止这种潜在的毒性积聚。
理解分隔效应的一种方法是检查它是如何影响系统中存在的守恒量的。在本例中,我们计算了两种模型中的守恒量t . brucei并在Bakker等人的分析背景下讨论它们的意义。
加载项目
首先使用命令行加载项目SBIOLADP项目.
SBIOLADP项目锥虫糖酵解
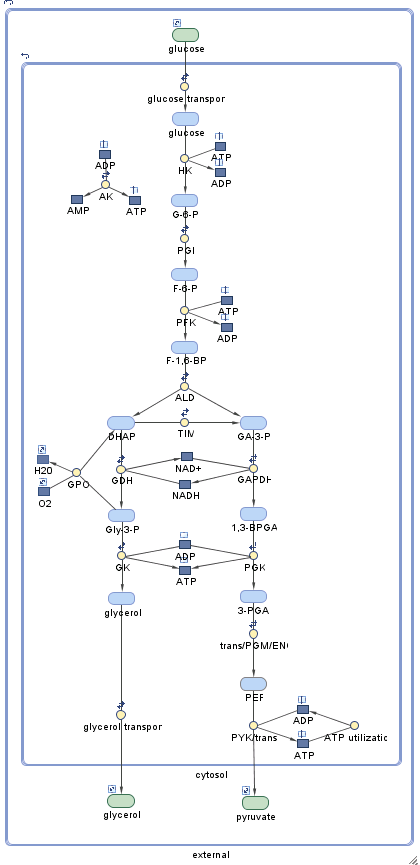
该项目包含两个模型。第一个模型,m1,包含如下所示的野生型糖酵解网络。(您可以通过启动SimBiology Model Builder应用程序来交互式地探索网络simBiologyModelBuilder打开位于(matlabroot/toolbox/simbio/simbiodemos.)的项目文件trypanosome_glycolysis.sbproj

这个系统是Bakker等人使用的途径的稍微简化版本。该模型有三个室:糖体、胞质和胞外。糖体中所含的代谢物为蓝色,而胞质内或胞外的代谢物为绿色。有些种类,如甘油3-磷酸(Gly-3-P),存在于多个室中。
这个通路开始于将细胞外葡萄糖导入糖体(为了方便起见,在这个过程中“跳过”细胞质)。在图中,这条通路向下进行,以丙酮酸的运输出胞质结束。在有氧条件下,甘油3-磷酸(Gly-3-P)被糖体外的甘油3-磷酸氧化酶(GPO)氧化;结果,葡萄糖完全转化为丙酮酸。在厌氧条件下,这个反应不发生,糖酵解途径产生甘油和丙酮酸。
计算野生型网络中的守恒量
这个函数SBIocons部分检查一个模型的化学计量矩阵的结构,以发现守恒物种的线性组合。这种分析是结构性的,因为它只依赖于网络的化学计量和结构,而不依赖反应动力学。事实上,这个模型中的所有反应速率都被设为0,因为这些速率对我们的分析并不重要。这里我们所说的SBIocons部分使用算法规范“semipos”,因此所有返回的守恒量只包含物种的正总和。第三个参数“p”要求将输出打印到字符串单元格数组中。
cons_wt = sbioconsmoiety (m1,“semipos”,“p”)
cons_wt =10 x1细胞{的外部。葡萄糖的}{的外部。甘油的}{的外部。丙酮酸的}{胞质。净水}{“胞质。O2的}{胞质。Gly-3-P +细胞溶质。DHAP}{“胞质。ATP +细胞溶质。ADP +细胞溶质。AMP的}{glycosome。ATP + glycosome。ADP + glycosome。AMP的}{glycosome。NAD + + glycosome。NADH'}{'2糖体。ATP + glycosome。ADP + glycosome。G-6-P + glycosome。f6 - p + 2糖体。f - 1, 6-BP + glycosome。DHAP + glycosome。GA-3-P + glycosome.Gly-3-P + glycosome.1,3-BPGA'}
单元格数组中的最后一个单元格包含一个长字符串。将此字符串拆分并显示,以便读取。
disp(cons_wt{end}(1:68));
2糖体。ATP+糖体。ADP+糖体。G-6-P+糖体。F-6-P
disp (cons_wt{}结束(69:147));
+2糖体。F-1,6-BP+糖体。DHAP+糖体。GA-3-P+糖体。Gly-3-P
disp (cons_wt{}结束(148);
+糖体1,3-BPGA
产量SBIocons部分包含10个量,其时间变化率为零,与反应动力学无关。腺嘌呤核苷酸ATP、ADP和AMP有两个保守的池,一个在糖体中,一个在细胞溶胶中。烟酰胺核苷酸NAD+和NADH的糖体池也是保守的。单独保守的物种如external.glucosoe和cytosol.O2是系统边界上的物种,其BoundaryCondition属性设置为true。这些物种包括在SBIocons部分因为在假设的模拟中,它们的数量确实会保持不变。
剩下的两个保守量代表结合的磷酸盐池,一个在糖体内部,一个在糖体外部。里面的一个包括9个不同的物种。注意,ATP和果糖-1,6-二磷酸(f -1,6- bp)的系数都是2,因为这些物种都有两个可转移的磷酸基。
以下是参与保护总和的物种。这个图是通过在SimBiology桌面的Diagram Table View中选择相关的物种而生成的。当葡萄糖被ATP磷酸化形成葡萄糖6-磷酸(G-6-P)时,保守循环“开始”。这个磷酸基通过通路向下传播,直到它从1,3-二磷酸甘油酸(1,3- bpga)或糖体甘油3-磷酸(Gly-3-P)转移回ATP,完成这个循环。

请注意细胞质总和。DHAP +细胞溶质。Gly-3-P作为一个独立的保守库出现,是因为DHAP/Gly-3-P逆向转运体将一个糖体DHAP分子与一个胞质Gly-3-P分子交换,反之亦然。有磷酸基的通量进出这个池,但下一个通量是零,因为这些通量相互抵消了。
查看无糖体的实验模型
现在让我们考虑第二个模型,平方米,其中包含生物信息学Bakker等人的实验网络,其中糖体已被去除。在该模型中,所有代谢物均存在于胞浆中。特别是,糖体内外不再存在DHAP和Gly-3-P的反转运交换,腺嘌呤核苷酸ATP、ADP和AMP存在单一池。
计算实验网络中的守恒量
cons_exp=sbiocons部分(m2,“semipos”,“p”)
cons_exp =7x1电池{的外部。葡萄糖的}{的外部。甘油的}{的外部。丙酮酸的}{胞质。净水}{“胞质。O2的}{胞质。NADH +细胞溶质。NAD + '}{胞质。AMP +细胞溶质。ADP +细胞溶质。ATP的}
系统边界上的物种在实验模型中仍然存在,它们的数量再次守恒。然而,没有了糖体,结合的磷酸盐的守恒就消失了,只留下烟酰胺和腺嘌呤核苷酸的守恒关系。
讨论
在他们分析糖体的功能t . brucei, Bakker等人发现,糖体区隔阻止了糖酵解过程中己糖磷酸中间体G-6-P和f -1,6- bp的潜在毒性积累。根据观察到的有糖体和没有糖体的磷酸盐在保存方面的差异,可以理解这种观察。当糖体存在时,中间产物如G-6-P或f -1,6- bp不能积聚到任意高水平,因为它们受到保存池中存在的有机磷酸盐总量的限制。如果没有糖体,就没有这种限制。也可以通过考虑腺嘌呤核苷酸的糖体区隔来获得见解。当细胞外葡萄糖水平升高时,刺激HK和PFK反应。当糖体存在时,这些反应是自我限制的,因为它们从糖体ATP、ADP和AMP的保守池中耗尽ATP。另一方面,当糖体不存在时,胞质ATP/ADP比率实际上随着细胞外葡萄糖水平的增加而增加。因此,HK和PFK的反应被进一步刺激,导致它们的产物G-6-P和f -1,6- bp的积累。下载188bet金宝搏
这一分析表明,糖体区隔对中间体的形成提供了一个负反馈机制。Bakker等人认为,有机磷酸盐的保存池也可以作为野生型的能量储存机制t . brucei在饥饿的时候。
在这个例子中,我们展示了如何在SimBiology模型中计算守恒量,以及如何通过分析这些守恒量来洞察网络的行为。
参考文献
dr . Bakker, B. M., Mensonides, F. I. C., Teusink, B., van Hoek, P., Michels, P. A. M., and Westerhoff, h.v.区隔保护锥虫免受糖酵解的危险设计。PNAS (2000) vol. 97, 2087-2092。
Bakker,B.M.,Michels,P.A.M.,Opperdoes,F.R.,和Westerhoff,H.V.血液形式的糖酵解布氏锥虫可以理解在动力学的糖酵解酶。生物。化学。(1997)第272卷,327 -3215页。
您还可以从以下列表中选择网站: